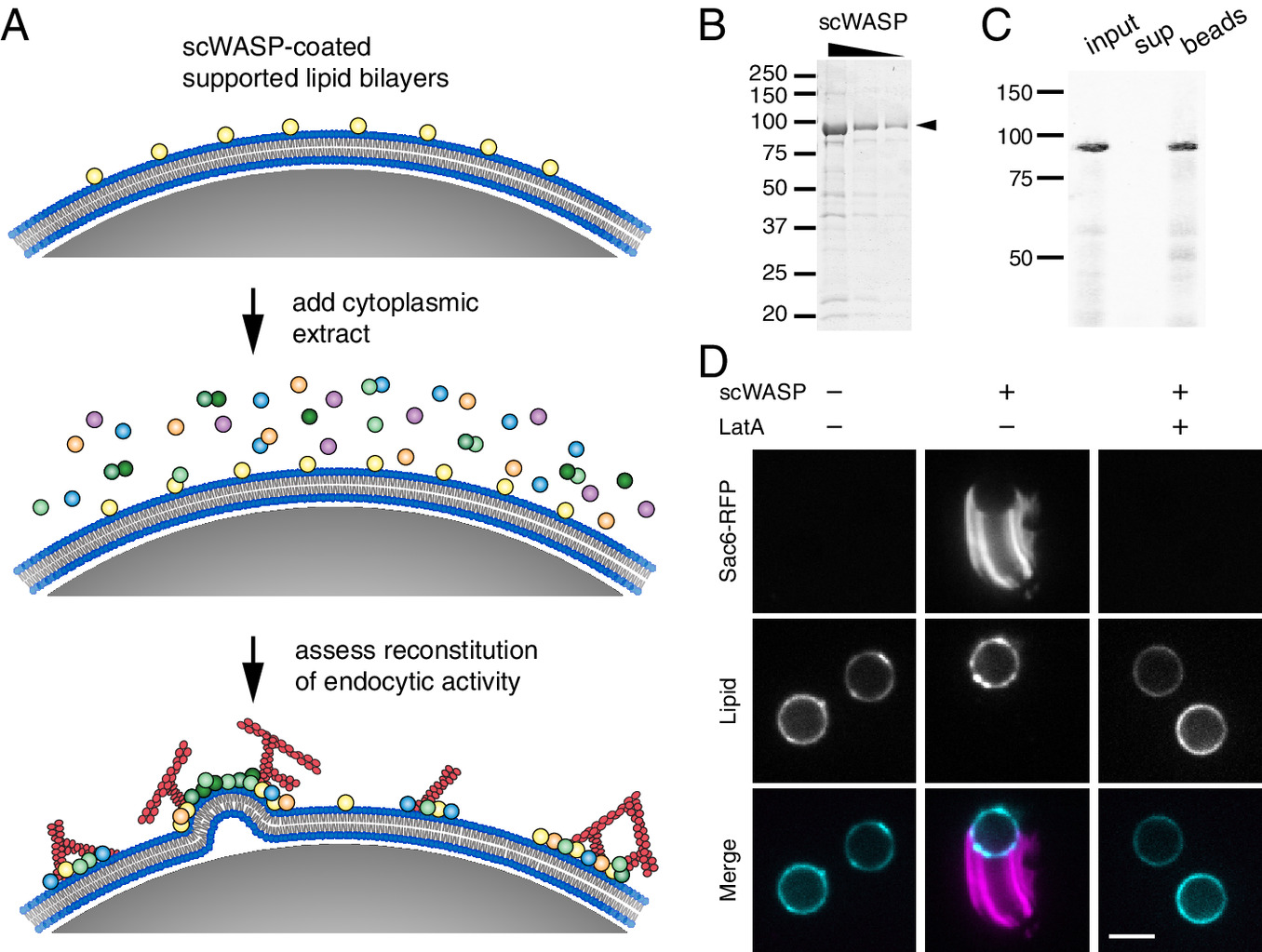
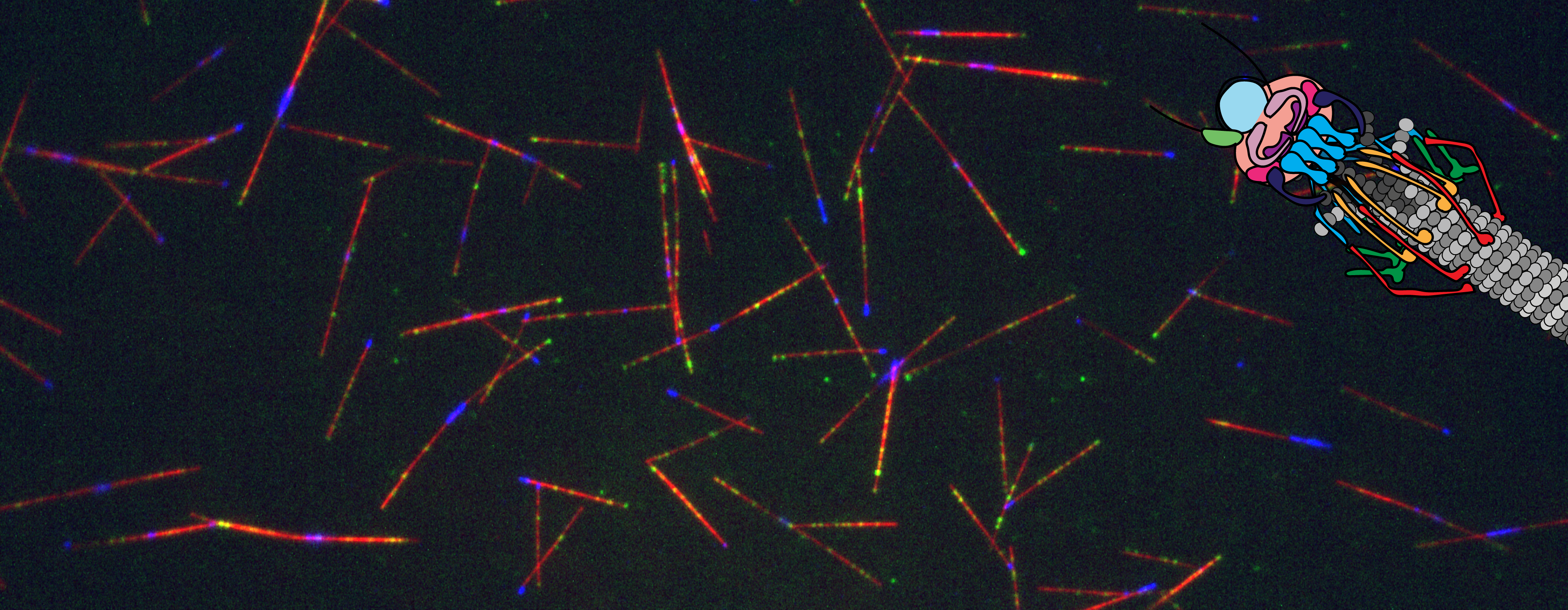
One of the saddest parts of being in a lab, is we become family, but yet people do have to move on with their lives. On one hand, it’s great we end up with friends sprinkled all around the world, but sometimes you just really miss them. And since the last website update, both Zane and Meiyan have moved on to new adventures. Truly, I am so happy for them. Zane and his family have moved down to San Diego were he now is a staff scientist in the Oegema/Desai lab. Finally, his kids can wear shorts all year and have it not being weird and cold! Meiyan and her family are also in warmer climates as they have moved down to Florida, specifically the University of Florida, where her and her husband will continue their research! We all wish them luck, happiness, and success in their new adventures.

The lab gives a farewell toast to Zane.

David says goodbye to Meiyan.