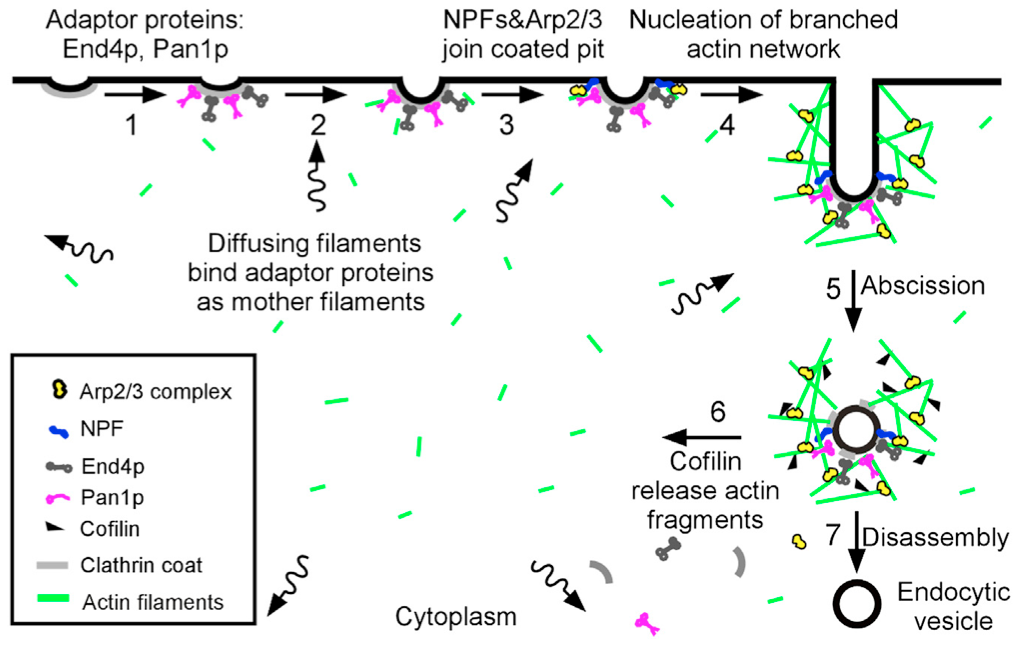
“Sever, diffuse and trigger” model for actin filament turnover in actin patches. The 7 steps (numbers next to the arrows) are (1) clathrin coated pits bind adaptor proteins End4p and Pan1p, (2) short, diffusing actin filaments bind to End4p and Pan1p associated with coated pits, and (3) Arp2/3 complex interacts with these mother filaments and nucleation promoting factors to (4) initiate branching nucleation of actin filaments that promote elongation of the endocytic tubule. (5) After abscission of the vesicle, (6) cofilin severs actin filaments to generate a pool of short, diffusing actin filaments, some of which return to the cycle at step 2.
Schematics illustrating the mathematical model. Shown are the Cdc42 autoamplification module (A), the complete endocytosis module (B), the exocytosis module (C), and a legend for graphics (D).
Here’s the story of a lovely lady.
Who studied what a microtubule’s for.
All her group loves mitosis, like Georjana,
And kinetochores.
Here’s the story of a PI named David,
Who was interested in actin in live cells,
And his group mapped endocytosis dynamics,
Yet they were by themselves.
After their post-docs, where the lady met this fellow,
And named a protein complex DAM instead of DARN,
They knew their groups must form one laboratory.
That’s the way our lab became the Drubin/Barnes!
The Drubin/Barnes!
PtdIns(4,5)P2-positive intracellular vesicles, visualized with GFP- or RFP-PHPLCδ, colocalize with endocytic clathrin machinery components (endogenous AP-2, epsin 1, and dynamin 2–GFP) but not with markers of endosomes (endogenous EEA1, GFP-Rab5, and GFP-Rab9), the endosomal phosphoinositide PtdIns3P (GFP-2×FYVEHrs), and Golgi-localized PtdIns4P (GFP-PHOSBP). For clarity, PHPLCδ is false colored green, and the second marker is false colored red in all images.
Yidi Sun‘s new paper is out now as an electronic publication ahead of print in the Journal of Cell Science. Congratulations to Yidi on her great work! The abstract is below. The PDF can be downloaded from JCS here.
Anionic phospholipids PI(4,5)P(2) and phosphatidylserine (PS) are enriched in the cytosolic leaflet of the plasma membrane where endocytic sites form. In this study, we investigated the roles of PI(4,5)P(2) and PS in clathrin-mediated endocytosis (CME) site initiation and vesicle formation in Saccharomyces cerevisiae. Live-cell imaging of endocytic protein dynamics in an mss4(ts) mutant, which has severely reduced PI(4,5)P(2) levels, revealed that PI(4,5)P(2) is required for endocytic membrane invagination but is less important for endocytic site initiation. We also demonstrated that in various deletion mutants of genes encoding components of the Rcy1-Ypt31/32 GTPase pathway, endocytic proteins dynamically assemble not only on the plasma membrane but also on intracellular membrane compartments, which are likely derived from early endosomes. In rcy1Δ cells, fluorescent biosensors indicated that PI(4,5)P(2) only localized to the plasma membrane while PS localized to both the plasma membrane and intracellular membranes. Furthermore, we found that polarized endocytic patch establishment is defective in the PS-deficient cho1Δ mutant. We propose that PS is important for directing endocytic proteins to the plasma membrane and that PI(4,5)P(2) is required to facilitate endocytic membrane invagination.
Genetic Engineering & Biotechnology News is out with a Feature Article this week including some comments from David Drubin about targeted genome modification in mammalian cells for fluorescence microscopy studies.
“The difference between taking snapshots of the process and watching a movie is just night and day,” says David Drubin, Ph.D., professor of cell and developmental biology at the University of California, Berkeley, whose lab uses fluorescence to understand the intricate details underlying clathrin-mediated endocytosis.
Researchers in David Drubin’s lab at the University of California, Berkeley genetically engineered a human cell line to express endogenous levels of RFP-tagged clathrin light chain A (red) and GFP-tagged dynamin 2 (green) for studying clathrin-mediated endocytosis. The above 3D kymograph of the cell surface, with the time dimension in the z-axis, shows the full lifetime of hundreds of clathrin patches on the membrane, which terminate upon recruitment of dynamin. [Aaron T. Cheng]
The May 2012 edition of Biowire, a publication of Sigma-Aldrich, includes an interview with David Drubin about the projects in our lab looking at clathrin-mediated endocytosis (CME) in mammalian cells using zing finger nuclease (ZFN) technology to undertake targeted genome modification. Traditionally, CME has been studied in cells in which fluorescently-tagged components of endocytic machinery are overexpressed using exogenous constructs. Data obtained in many labs using these methods suggested that CME was highly variable. Using ZFN technology, in collaboration with Sangamo Biosciences, our lab recently showed that CME is robust and efficient in mammalian cells. The new results highlight the technical advantages of tagging genes at their endogenous loci, an approach that has been historically limited to genetically tractable organisms, such as the Drubin/Barnes Lab favorite Saccharomycescerevisiae (budding yeast). Emerging technologies, such as ZFNs and TALENs, however, are now making this sort of precise genomic manipulation possible in animal cells, including human cells, giving us new and powerful ways of studying cellular biology.
Cellular processes should be studied as close to their natural states as possible. I suspect that, as we see more uses of zinc finger nucleases [for tagging endogenous genes], people will find that they have been inadvertently perturbing the processes that they have been studying.